유전"이란? 이라는 간단한 질문에 대한 개인의 의견과 "유전" 현상의 의미
런 이해가 자연스럽게 "어떻게 일어나는 거지?" 라는 과학적 질문으로 이행되고 있었습니다. 이러한 호기심은 여러분들이 학습하는데 매우 중요한 동기가 될 것이며, 강의를 통해 조금은 이해 할 수 있을 것입니다. 하지만, 유전학은 현재 진행형인 학문입니다.
다시 말해, 현재 우리는 유전현상의 원인과 그 메카니즘에 대해 궁극적으로 알 지 못 한다는 것이며. 이런 측면에서 "어떻게 유전학을 연구하는지"에 대한 학습도 중요한 포인트가 될 것입니다.”유전학의 이해“를 통해 유전학연구방법도 접해 볼 수 있는 기회가 되었으면 합니다.
우선 유전은 수많은 담고 있고 이것이 세대를 거치면서 형질과 생물학적 정보가 전이되고, 이것이 복잡하게 하나 이상의 유전자와 환경요인에 의해서 결정된다고 한다.
그리고 유전형질에는 집단에는 수많은 대립유전자가 있고, 대립유전자가 높은 빈도로 나타난다고 한다. 이것을 통해 집단을 이해하기 위해서 는 유전을 분석하는 것이 의미 있다고 생각이 된다.
그러므로 유전은 생명과학의 중심으로, 각종 질병 등의 문제해결에 열쇠가 되는 분야라고 생각한다. 그리고 질병을 해결하기 위해서는 유전자 발현의 추적을 통해 우리는 질병에 대한 새로운 정보를 밝혀낼 수 있을 것이라고 생각한다
우리 몸은 동일한 유전정보를 가진 서로다른 세포들로 구성되어 있다. 이들 세포들의 정상적인 기능은 인체의 항상성을 유지하는데 있어 매우 중요하다. 정상적인 세포기능은 세포 내 다양한 소기관들의 유기적인 상호작용을 통해 이루어지며, Chap2에서는 이들 세포의 특징에 대해 다루고 있다.
우리는 유전정보의 작용에 의해 겉으로 표현되는 모습 (phenoypes or traits)을 바탕으로 유전현상을 이해하고 있습니다. 특정 개체가 유전정보를 표현하는 주요 방법은 세포의 기능조절입니다. 이를 통해 다양한 traits의 출현이 가능합니다. 우리가 쉽게 이들을 인지할 수 있는 대표적인 traits은 유전질환입니다. 이들의 거시적인 표현형도 세부적으로 들여다보면, 세포기능 변화가 그 주요원인인 경우가 대부분입니다. Chap2에서는 이러한 예시들을 통해 세포의 기본적인 기능과 표현형의 상관관계를 이해 할 수 있습니다.
세포소기관들은 다양한 기능을 수행하고 있다. 세포는 이들에 의
해 유전정보의 보관과 사용, 에너지 획득, 물질의 분비, 그리고 잔해
를 분해한다. 소낭 같은 세포소기관은 다른 세포 구성물에 해를 끼
칠 수 있는 생화학물질을 격리시키고 있다. 어떤 세포소기관은 특정 |
분자를 만드는 화학반응에 참이하는 효소가 순시에 맞춰 점점이 박
혀 있는 막을 지니고 있다.
대부분의 세포에서 가장 두드리진 세포소기관은 핵이다. 핵막이라
고 하는 층이 핵을 둘러싸고 있다. 마치 선박 양쪽에 나 있는 둥근 창
과 같이 구멍 주위에 단백질의 고리들을 지닌 핵공을 통해 핵에서의
생화학물질들의 출입이 이루어진다
대부분의 세포에서 가장 두드러진 세포소기관은 핵이다. 핵막이라.
| 고 하는 층이 핵을 둘러싸고 있다. 마치 선박 양쪽에 나 있는 둥근 창
과 같이 구멍 주위에 단백질의 고리들을 지닌 핵공을 통해 책에서의
생화학물질들의 출입이 이루어진다(그림 2. 5).
핵막의 안쪽 면에는 핵막하층(nuclear lamina)이라 불리는 한 충
의 섬유성 물질이 있다. 이 층은 몇 가지 중요한 기능을 한다. 핵 안
의 DNA는 세포분열의 신호로 핵막하층과 접촉한다
. 또한 핵막하층
은 핵공을 제자리에 붙잡는 기계적인 지지대로 작용한다. 제3장의 서
두에서 논의하는 이주 드물지만 가속화된 노화 장애가 비징상적인
핵막하층에 의해 생긴다.
현미경으로 관찰하면 핵 안쪽에 조금 어둡게 보이는 부분이 있는
| 데 이것이 인(nucleolus, 작은 핵')이고, 이기시 리보솜이 만들이진
다.
해에는 DNA가 많은 단백질들과 엉킨 형대의 연색체가 들어 있
| 다. 다른 단백질들은 섬유 형태로 핵안을 재위 핵이 기의 구형의 모양
을 유지하게 한다. RNA도 역시 많이 있을 뿐 아니라, DNA로부터
| RNA를 합성하는 데 필요한 단백질과 효소들도 들이 있다. 이들을 제
위한 핵 내의 액상 성분을 핵질(nucleoplasm)이라 부른다.
| 핵, 세포소기관, 맨 바깥쪽의 원형질막(plasma membrance)을 제외한 세포의 나머지 성분을 세포질(cytoplasm)이라 한다
. 세포 내다른 성분들로는 저장성인 단백질, 탄수화물, 지질, 그리고 색소 분
자와 여러 다른 작은 화학물질들이 있다. 이들을 제외한 세포질을
세포기질(cytosol)이라 한다.
세포소기관은 기본적인 생명 기능을 조절하고 특화된 세포 형태로의 특징을 제공하기 위해 서로 협조한다.
새포로부터 물질을 내보내는 분비과정은 세포소기관들의 기능 협조체계를 잘 보여준다
세포소기관들이 세포의 특수 기능율 제공한다 이들 구조는 대부분 투명하지만 여기에서는 색깔을 달리하였다
아파트에 쓰레기가 흩어져 있듯이 세포 안에도 온갖 쓰레기들이 생기
난다. 리소좀(ly sosome, 자르는 몸)이라 불리는 세포소기관이 쓰
레기를 처리한다. 리소좀은 막으로 둘러싸인 주머니로 그 속에 세균
잔해, 폐기된 세포소기관, 과다한 콜레스테롤과 같은 물질들을 분해
할 수 있는 효소를 함유하고 있다(그림 2.7), 이 효소는 어떤 소화된
영양물질을 세포가 사용할 수 있는 형태로 분해한다.
리소좀은 세포 안팎의 쓰레기들을 지니고 있는 소포와 융합한 후
리소 효소를 이용하여 그 내용물을 분해한다. 세포가 자신의 쓰레
기를 없애버리는 과정을 자신을 먹기'라는 의미의 자가소화작용(au-
tophagy)이라 부른다. 예를 들어 엔도솜(endosome)이라 불리는
원형질막에서 생긴 일종의 소낭은 과다한 낮은 밀도 리포단백질
(low-density lipoprotein, LDL) 콜레스테롤을 리소좀으로 데려간
다. 채워진 리소좀은 원형질막 쪽으로 이동하여 막과 융합하고 그 안
의 내용물을 밖으로 내보낸다. 리소좀은 자신의 효소가 활성을 유지하
기 위해 강한 산성을 띠지만, 다른 세포성분에는 피해를 주지 않는다.
세포마다 지니고 있는 리소좀의 숫자가 다르다. 온몸을 떠돌며 세
균을 먹어치우는 혈액세포의 어떤 것과 대식세포에는 리소좀이 굉장
히 많다. 콜레스테롤, 독성물질, 약품 등을 분해하기 위해 간의 세포
에도 리소좀이 많이 들어 있다.
모든 리소좀에는 43종류의 소화효소가 들어 있는데 이것들은 정확
한 균형이 유지되어야만 한다.
페르옥시솜(peroxisome)은 외막에 이러 가지 효소가 부착되어 있
는 주머니이다. 이 효소들은 다양한 기능을 수행한다. 특정 지질과
회귀한 화학물질의 분해, 지방 소화에 사용되는 담즙산의 합성, 그리
고 활성산소 노출에 의해 생긴 물질을 해독한다. 페르옥시행은 독성
물질을 다루는 간과 콩팥 세포에 많이 들어 있다.
에너지 생산- 미토콘드리아
세포가 생명의 화학반응을 하려면 지속적인 에너지가 필요하다. 음
식물에 있던 영양 분자의 화학 결합을 파괴하여 에너지를 나오게 하
는 것이 세포소기관인 미토콘드리아(mitochondria)이다.
미토콘드리아의 외막은 소포체나 골지체의 것과 비슷하며, 내막은
크리스테(cristae)라고 하는 주름진 구조를 하고 있다(그림 2.8), 이
크리스테에는 효소가 부착되어 있어 영양 분자로부터 에너지를 뽑아
내는 생화학반응을 촉매한다. 뽑아낸 에너지는 아데노신 삼인산
(adenosine triphosphate, ATP) 분자의 결합에너지로 보관된다.
이 ATP는 세포가 에너지를 필요로 할 때 마치 에너지 직불카드처럼
사용된다.
| 세포에 들어 있는 미토콘드리아의 수는 그 세포의 활동 양상에 따
라 몇백 개에서 수만 개로 매우 다양하다.
원형질막은 세포 간소통을 가능하게 함
원형질막에돌출한단백질. 당단백질. 당지질 등은 다른 1I포와의 세 포간 상호작용에 중요한표면 지형을 만든다
. 당신 세포의 표면은 그 자체가 당신 몸의일부분일 뿐 아니라 특정 기관이나 조직형의 한 부분이다. 원형질막에 부착되어 있는 많은 분자들은 수용체로 세포 밖의 분 자에 딱 맞71결합할 수 있는 모양이나 요면구조를 ㅎ하고 있다. 수용 체에 결합하는 분자를리간드라 하는데,이로써 분열과 같은 특별 각 용을 하는 세포 내에서화하1반응이 계속 이어져 일어날 수 있다.
세포 사이의 대화 과정을 신호전달 (signal transduction)이라 한 다.
. 세포접착 (cellular adhesion)이란 과정에서 원형질막이 세포를 어떤 다른 세 포들과 접착하도록 도와준다.이들 세포와 세포 시이의 대화는 조직을 이루는 데 중요하다.
잘못된 신호전달이나체포접착은 건강에 위험하다.
세포골격을 유지하는 기본요소들은 미세소관(microtubule) 미세섬유(microfilament). 그리고 중간섬유 intemeduatefilament)의 게 세가지이다(그림2.11) 이들은 단백질의 형태. 직경. 그리고 어 게 이들이 합쳐I서보다 더 큰 구조로되느냐에 구별된다. 이에 다른 단백질들이 결합하여 골격을 형성하여 세포가 외부의힘에 l리지 않고 특유의 모양을 유지하게 하는 힘을제공한다.
있다. 세포는 형성된 미세소관지 개개의 뒤불린 분자들을 모두 지니고 있다. 예를들이세포분열에서처럼미세미소관의 특수한 기능이 필요해지면 튜블린 이량채는 서로 조합하여 v1세소관을 형성한다.
분열이 끝 나면 일부 미세소관은다시 불린린 이량체로 해제되어 자후 작업의 원 재료로 보충된다. 세포는 끊임없이 미세소관을 조립하고 분해한다 미세소관은 머리카락과 같은 구조의 섬모(cilia, 라틴어로 '세포의 속눈썹'이란 의미)를 형성한다(1림 2.12).
섬모에는 움직이는 운동 섬모와 움직이지 않고 감각 기능을 지닌 일차심모의두 가지 형태가 운동섬모는 일차 미세소관보다 하나 이상의 쌍으로 된 미세소관 올 갖고 있다.
운동섬모 조화를 이문 이동은물결을 일으켜 세포 자체를 이동하게 하거나표면을 따라 물질을나아가게한다
.
Chap2전반부는 세포의 기본적인 구조와 그 소기관에 대한 내용이고, 후반부는 세포의 기능 중 가장 중요한 세포분열조절에 대한 내용입니다.
특히, 중요하게 봐야 할 부분은 cell cycle regulation입니다. 기본적으로 세포는 정상적인 세포분열을 위해 "cell cycle checkpoint"를 거치게 되고, 이러한 checkpoint 시점을 통과하지 못하면 세포는 사멸하게 됩니다.
G0, G1, S, G2 , M phase에 대한 설명과, cell cycle checkpoint의 종류 및 그 내용에 대한 설명을 과제로 제출하면 됩니다.
세포분열은 발생, 성장, 노화 및 생체 항상성 유지를 가능하게 하는 기본적인, 그리고 근본적인 생명활동입니다.
당연히, 세포분열은 정교한 과정을 통해 조절되며 (세포 분열을 유도하는 다양한 외부자극, 외부자극을 수용하는 다양한 신호전달 체계, 세포 분열 과정을 조절하는 시스템, 세포분열이 정상적으로 작동하는 지 점검하는 시스템, 세포 분열에 문제가 있을 때 Apoptosis를 유도하는 시스템 등으로 구성됨) 세포분열 결과는 다양한 phenotypes (traits)으로 표현됩니다.
세포분열 과정은 세포생물학에서 주로 다루는 주제이며, 유전학에서는 다양한 phenotypes을 세포 수준에서 이해 할 수 있도록 배경지식으로 활용 될 것입니다.
사람의 몸에서 조직에 따라 서로 다른 비율로 새로운 세포가 생기면
그에 맞춰 오래된 세포는 죽는다. 성장, 발달, 건강 유지와 질병이나
상처로부터의 치유는 DNA를 나누는 유사분열(mitosis)과 나머지
세포를 나누는 세포질분열(catokincesis)과 죽음의 한 형태인 세포자살
(apoptosis)의 속도 사이의 복잡한 상호작용이 요구된다.
간기 왕성한 활동의 시기
간기 동안에 세포는 생명의 기본적인 생화학적 기능을 계속하는 한편
DNA나 기다 세포내 구조 성분들을 복제한다. 간기는 2개의 간극기
인 G,기(G, phase)와 GI(G, phase), 그리고 하나의 S 합성기(S
phase)로 구분된다
세포는 활동이 없는G0기로 들어가기 위해,
G1기의 세포주기에서 따져나올 수 있다. G0 기의 세포는 세포의 특별한 특징
은 유지하지만 DNA의 복제나 분연은 하지 않는다. G0 기로부터 세포는 다시 유사분열을 계속 진생할 수도 있고, 혹은 죽기도 한다. 만약 암이 생길 수 있을 정도로 세포의 DNA 손상이 너무 크면세포자살로 간다. 결국 G0 기는 세포의 운명이 결정되거나 보류되는 시기이다.
유사분열 다음에 오는 G1,기 동안에 세포는 단백질, 지질, 그리고 탄수화물의 합성을 계속한다.
이 분자들은 세포분열의 결과로 생길 2개의 새로운 세포의 원형질막을 구성하는 데 사용될 것이다.
세포의 종류에 따라 세포주기의 지속 시간에서 가장 차이가 많은 것이 G1기이다. 간에 있는 세포처럼 느리게 분열하는 세포들은 G0기에서 빠져나와
수년 동안 G1기에 있기도 한다. 반면에 골수에 있는 빠르게 분열하는 세포들은 G1기를 16-24시간만에 끝낸다. 초기 배아의 세포에서는 G, 기가 생
략된다.
S기 동안에 세포는 유전체 모두를 복제한다. 그결과, 각각의 염색체는 2배로 늘어난 상태로 동원체(centromere) 부위에서 접합되어 있다. 대부분의 사람 세포들은 S기를 8~10시간에 끝낸다. 많은 단백질들이 또한 이 시기에 합성되는데이 중에는 염색체를 끌어당기 분리시키는 유사분열 방추체(spindle)
를 형성할 단백질들도 있다. 미세소관은 핵 가까이에 중심립(centn-ole)이라는 구조를 형성한다. 중심림 미세소관은 다른 단백질과 결합하고, 서로 직각방향으로 쌍을 이루어 장방형의 구조인 중심체(cen-tresome)를 만들고, 이는 방추체 내로 다른 미세소관을 배열한다.
G2, 기는 DNA가 복제되고 난 후 하지만 유사분열이 시작되기 전에 일어난다. 보다 많은 단백질들이 이 시기 동안에 합성된다. G1,기에 만들어진 분자들로부터 막이 조립되어 작고, 속이 빈 소포의 형태로원형질막 바로 밑에 보관된다. 이 소포들은 합쳐져서 딸세포를 둘러싸는 원형질막으로 될 것이다.
전기 (prophise)는 유사분열의 첫 단계로 DNA가 단단히 꼬인다.
이로씨 염색체는 보다 쉬게 분리된 수 있도록 잡고 두꺼워진다 (그리
2 16) 세포질에서는 불린을 이용한 미세소관이 형성되어 방추사
를 준비한다. 전기가 끝나갈 무렵 핵막이 소실된다. 인은 더 이상 보
이지 않는다.
중기(mmetaplie)는 전기 다음에 오는데, 방추사가 임색세의 동원
제 부위에 연결되며 염색체는 세포 중앙인 적도면에 정리한다. 중기
염색세는 큰 장력 하에 있지만 양쪽에서 당기주는 힘이 같아 움직임이
없는 것처리 보이는데 이는 마치 중다리기 밧줄이 생방하게 당겨지는
것과 같다.
다음으로 후기 (anaphase)가 진행되면시, 중기 염색제가 정립한 중
양부에서 원형질막의 만입이 이루어진다. 원형질막 안쪽 면에 미세섬
유가 띠를 형성하면서 세포를 양분되게 조여들이 간다. 동인체가 같
라지면서 장력은 사라지고 쌍을 이루고 있던 염색분제는 세포의 양
복으로 하나 이동되는데, 마치 줄다리기를 하다가 줄이 끊어지면
사람들이 양쪽으로 쓰러지는 것과 같다. 미세소관의 이동은 분인하
는 세포를 더 멀어 나가게 한다. 아주 짧은 후기 동안에, 세포는 잠
간 정상보다 2배인 협색세를 지닌 상대가 된다. 왜냐하면 각각의 염
색분체는 독자적인 염색세가 되었지만 세포는 아직 물리적으로 분리
되지 않았기 때문이다.
말기(telophise)는 유사분열의 마지막 단계로, 세포는 양에 한
세트씩의 염색제들을 가진 아처럼 보인다. 길게 늘어난 세포의 각
끝에서 방추사는 소실되고, 인과 책 주위의 막은 재생된다. 유전물
의 분열은 완료된 상대이다. 뒤이어 세포질분열(cytokinesis)이 진행
되어 세포소기관과 고분자들이 2개의 딸세포로 나뉘어 들어간다. 마
지막으로 미세섬유 때는 졸라매는 끈처럼 수축하이 새로 생긴 2개의
세포로 분리시킨다.
상호작용을 하는 단백진 집단이 체크포인트라 불리는 세포주기의
특징 시기에 작용하이 염색제들이 정확하게 복제되고 그리고 딸세포
로 나뉘어 들어가도록 한다(그림 2. 17). 예를 들어 'DNA 손상 체크
포인트'는 특정 단백질들이 손상된 DNA를 회복할 때까지 일시적으
로 세포주기의 진행을 징지시킨다. 세포자살 체크포인트'는 유사분
연의 시작에 좌우된다.
이 체크포인트 동안에 서바이빈 (sunivin)이란 단백질은 세포로 하여금 죽으라는 신호를 무시하게 하여 그 결과죽으라는 세포자살을 하지 않고 분열하라는 유사분열을 하게 한다.세포들은 자기에게 남은 분열 횟수를 어떻게 알 수 있을까?
그 해 답은 염색체의 끝부분인 말단소체(telomer)에 있다(그림 2 18), 만
단소체는 말단부터 잃게 되는 절편으로 태워버리는 세포의 퓨즈와 같은 기능을 한다. 말단소체 부분에는 6개의 특징 DNA 염기서열이 수백에서 수천 번 반복해서 배열되어 있다. 매 유사분열 때마다 50~200개의 가장 끝 부위의 염기들이 소실되어 염색체 크기는 점점 작아지게된다. 약 50회 정도의 분열을 하고 난 후에는 말단소체 DNA의 임계길이를 잃어버리게 되어 유사분열의 신호가 멈춘다. 세포는 더 이상분열 없이 생존해 있거나 죽게 된다.
호르몬과 성장인자들은 세포주기를 조절하는 외부의 화학신호들이다. 호르몬(homone)은 특정 샘에서 만들어져 힘액을 타고 신제의 다른 부위로 이동한 후 그곳에서 특별한 효과를 발휘한다. 예를들어뇌에서분비되는호르몬들은 여성 자궁에 배열하고 있는 세포에신호를 주어 매달 유사분열로 세포층을 쌓아 올려 임신이 가능하도록 준비를 하게 한다. 생장인자의 활동은 보다 국소적이다. 예를 들어 상피성장인자 (epidermal growth factor, EGF)는 상처 딱지 아래에 있는 피부의 세포분열을 자극한다. 어떤 암 치료약 중에는 암세포에 있는 성장인자 수용체를 막음으로써 분열하는 신호를 차단하도록 하는 작용을 이용한 것도 있다.
사이클린과 키나아제의 두 종류의 단백질은 세포 내에서 상호작용을 하여 그 산물이 유사분열을 하게 하는 유전자를 활성화시킨다.
두 단백질은 쌍을 이룬다. 사이클린의 환성은 세포주기 동안에 규칙으로 변화하는 반면 키나아제의 활성은 일정하게 유지된다. 일정 수의 사이클린 키나아제 쌍은 유사분열을 개시하게 하는 유전자를현시킨다. 그러고는 유사분열이 시작되면 효소는 사이클린을 분해
다. 다음 간기 동안에 사이클린이 복구되면 주기는 다시 시작된다.
Chap2전반부는 세포의 기본적인 구조와 그 소기관에 대한 내용이고, 후반부는 세포의 기능 중 가장 중요한 세포분열조절에 대한 내용입니다.
특히, 중요하게 봐야 할 부분은 cell cycle regulation입니다. 기본적으로 세포는 정상적인 세포분열을 위해 "cell cycle checkpoint"를 거치게 되고, 이러한 checkpoint 시점을 통과하지 못하면 세포는 사멸하게 됩니다.
G0, G1, S, G2 , M phase에 대한 설명과, cell cycle checkpoint의 종류 및 그 내용에 대한 설명을 과제로 제출하면 됩니다.
세포분열은 발생, 성장, 노화 및 생체 항상성 유지를 가능하게 하는 기본적인, 그리고 근본적인 생명활동입니다.
당연히, 세포분열은 정교한 과정을 통해 조절되며 (세포 분열을 유도하는 다양한 외부자극, 외부자극을 수용하는 다양한 신호전달 체계, 세포 분열 과정을 조절하는 시스템, 세포분열이 정상적으로 작동하는 지 점검하는 시스템, 세포 분열에 문제가 있을 때 Apoptosis를 유도하는 시스템 등으로 구성됨) 세포분열 결과는 다양한 phenotypes (traits)으로 표현됩니다.
세포분열 과정은 세포생물학에서 주로 다루는 주제이며, 유전학에서는 다양한 phenotypes을 세포 수준에서 이해 할 수 있도록 배경지식으로 활용 될 것입니다.
사람의 몸에서 조직에 따라 서로 다른 비율로 새로운 세포가 생기면
그에 맞춰 오래된 세포는 죽는다. 성장, 발달, 건강 유지와 질병이나
상처로부터의 치유는 DNA를 나누는 유사분열(mitosis)과 나머지
세포를 나누는 세포질분열(catokincesis)과 죽음의 한 형태인 세포자살
(apoptosis)의 속도 사이의 복잡한 상호작용이 요구된다.
간기 왕성한 활동의 시기
간기 동안에 세포는 생명의 기본적인 생화학적 기능을 계속하는 한편
DNA나 기다 세포내 구조 성분들을 복제한다. 간기는 2개의 간극기
인 G,기(G, phase)와 GI(G, phase), 그리고 하나의 S 합성기(S
phase)로 구분된다
세포는 활동이 없는G0기로 들어가기 위해,
G1기의 세포주기에서 따져나올 수 있다. G0 기의 세포는 세포의 특별한 특징
은 유지하지만 DNA의 복제나 분연은 하지 않는다. G0 기로부터 세포는 다시 유사분열을 계속 진생할 수도 있고, 혹은 죽기도 한다. 만약 암이 생길 수 있을 정도로 세포의 DNA 손상이 너무 크면세포자살로 간다. 결국 G0 기는 세포의 운명이 결정되거나 보류되는 시기이다.
유사분열 다음에 오는 G1,기 동안에 세포는 단백질, 지질, 그리고 탄수화물의 합성을 계속한다.
이 분자들은 세포분열의 결과로 생길 2개의 새로운 세포의 원형질막을 구성하는 데 사용될 것이다.
세포의 종류에 따라 세포주기의 지속 시간에서 가장 차이가 많은 것이 G1기이다. 간에 있는 세포처럼 느리게 분열하는 세포들은 G0기에서 빠져나와
수년 동안 G1기에 있기도 한다. 반면에 골수에 있는 빠르게 분열하는 세포들은 G1기를 16-24시간만에 끝낸다. 초기 배아의 세포에서는 G, 기가 생
략된다.
S기 동안에 세포는 유전체 모두를 복제한다. 그결과, 각각의 염색체는 2배로 늘어난 상태로 동원체(centromere) 부위에서 접합되어 있다. 대부분의 사람 세포들은 S기를 8~10시간에 끝낸다. 많은 단백질들이 또한 이 시기에 합성되는데이 중에는 염색체를 끌어당기 분리시키는 유사분열 방추체(spindle)
를 형성할 단백질들도 있다. 미세소관은 핵 가까이에 중심립(centn-ole)이라는 구조를 형성한다. 중심림 미세소관은 다른 단백질과 결합하고, 서로 직각방향으로 쌍을 이루어 장방형의 구조인 중심체(cen-tresome)를 만들고, 이는 방추체 내로 다른 미세소관을 배열한다.
G2, 기는 DNA가 복제되고 난 후 하지만 유사분열이 시작되기 전에 일어난다. 보다 많은 단백질들이 이 시기 동안에 합성된다. G1,기에 만들어진 분자들로부터 막이 조립되어 작고, 속이 빈 소포의 형태로원형질막 바로 밑에 보관된다. 이 소포들은 합쳐져서 딸세포를 둘러싸는 원형질막으로 될 것이다.
전기 (prophise)는 유사분열의 첫 단계로 DNA가 단단히 꼬인다.
이로씨 염색체는 보다 쉬게 분리된 수 있도록 잡고 두꺼워진다 (그리
2 16) 세포질에서는 불린을 이용한 미세소관이 형성되어 방추사
를 준비한다. 전기가 끝나갈 무렵 핵막이 소실된다. 인은 더 이상 보
이지 않는다.
중기(mmetaplie)는 전기 다음에 오는데, 방추사가 임색세의 동원
제 부위에 연결되며 염색체는 세포 중앙인 적도면에 정리한다. 중기
염색세는 큰 장력 하에 있지만 양쪽에서 당기주는 힘이 같아 움직임이
없는 것처리 보이는데 이는 마치 중다리기 밧줄이 생방하게 당겨지는
것과 같다.
다음으로 후기 (anaphase)가 진행되면시, 중기 염색제가 정립한 중
양부에서 원형질막의 만입이 이루어진다. 원형질막 안쪽 면에 미세섬
유가 띠를 형성하면서 세포를 양분되게 조여들이 간다. 동인체가 같
라지면서 장력은 사라지고 쌍을 이루고 있던 염색분제는 세포의 양
복으로 하나 이동되는데, 마치 줄다리기를 하다가 줄이 끊어지면
사람들이 양쪽으로 쓰러지는 것과 같다. 미세소관의 이동은 분인하
는 세포를 더 멀어 나가게 한다. 아주 짧은 후기 동안에, 세포는 잠
간 정상보다 2배인 협색세를 지닌 상대가 된다. 왜냐하면 각각의 염
색분체는 독자적인 염색세가 되었지만 세포는 아직 물리적으로 분리
되지 않았기 때문이다.
말기(telophise)는 유사분열의 마지막 단계로, 세포는 양에 한
세트씩의 염색제들을 가진 아처럼 보인다. 길게 늘어난 세포의 각
끝에서 방추사는 소실되고, 인과 책 주위의 막은 재생된다. 유전물
의 분열은 완료된 상대이다. 뒤이어 세포질분열(cytokinesis)이 진행
되어 세포소기관과 고분자들이 2개의 딸세포로 나뉘어 들어간다. 마
지막으로 미세섬유 때는 졸라매는 끈처럼 수축하이 새로 생긴 2개의
세포로 분리시킨다.
상호작용을 하는 단백진 집단이 체크포인트라 불리는 세포주기의
특징 시기에 작용하이 염색제들이 정확하게 복제되고 그리고 딸세포
로 나뉘어 들어가도록 한다(그림 2. 17). 예를 들어 'DNA 손상 체크
포인트'는 특정 단백질들이 손상된 DNA를 회복할 때까지 일시적으
로 세포주기의 진행을 징지시킨다. 세포자살 체크포인트'는 유사분
연의 시작에 좌우된다.
이 체크포인트 동안에 서바이빈 (sunivin)이란 단백질은 세포로 하여금 죽으라는 신호를 무시하게 하여 그 결과죽으라는 세포자살을 하지 않고 분열하라는 유사분열을 하게 한다.세포들은 자기에게 남은 분열 횟수를 어떻게 알 수 있을까?
그 해 답은 염색체의 끝부분인 말단소체(telomer)에 있다(그림 2 18), 만
단소체는 말단부터 잃게 되는 절편으로 태워버리는 세포의 퓨즈와 같은 기능을 한다. 말단소체 부분에는 6개의 특징 DNA 염기서열이 수백에서 수천 번 반복해서 배열되어 있다. 매 유사분열 때마다 50~200개의 가장 끝 부위의 염기들이 소실되어 염색체 크기는 점점 작아지게된다. 약 50회 정도의 분열을 하고 난 후에는 말단소체 DNA의 임계길이를 잃어버리게 되어 유사분열의 신호가 멈춘다. 세포는 더 이상분열 없이 생존해 있거나 죽게 된다.
호르몬과 성장인자들은 세포주기를 조절하는 외부의 화학신호들이다. 호르몬(homone)은 특정 샘에서 만들어져 힘액을 타고 신제의 다른 부위로 이동한 후 그곳에서 특별한 효과를 발휘한다. 예를들어뇌에서분비되는호르몬들은 여성 자궁에 배열하고 있는 세포에신호를 주어 매달 유사분열로 세포층을 쌓아 올려 임신이 가능하도록 준비를 하게 한다. 생장인자의 활동은 보다 국소적이다. 예를 들어 상피성장인자 (epidermal growth factor, EGF)는 상처 딱지 아래에 있는 피부의 세포분열을 자극한다. 어떤 암 치료약 중에는 암세포에 있는 성장인자 수용체를 막음으로써 분열하는 신호를 차단하도록 하는 작용을 이용한 것도 있다.
사이클린과 키나아제의 두 종류의 단백질은 세포 내에서 상호작용을 하여 그 산물이 유사분열을 하게 하는 유전자를 활성화시킨다.
두 단백질은 쌍을 이룬다. 사이클린의 환성은 세포주기 동안에 규칙으로 변화하는 반면 키나아제의 활성은 일정하게 유지된다. 일정 수의 사이클린 키나아제 쌍은 유사분열을 개시하게 하는 유전자를현시킨다. 그러고는 유사분열이 시작되면 효소는 사이클린을 분해
다. 다음 간기 동안에 사이클린이 복구되면 주기는 다시 시작된다.


내용은 유전자의 발현과 관련된 내용으로 주로 분자생물학 범주에 속하는 내용이기에 예년에는 대면강의 시 다루지 않던 부분입니다.
하지만 기본적으로 allele과 traits의 관계를 이해하기 위해서 필요한 배경지식입니다. 보통 일반생물학에서 다루던 내용으로도 현재는 충분하고, 이를 chap10, 11을 이용해 복습해 보면 됩니다.
우선, Chap10을 통해 유전자의 기본적인 구조를 이해하고 (Promoter, Enhancer, Intron, Exon etc..) 이들 각 구조의 기능을 정리해 보도록 하고, 유전자의 발현과정을 정리해 보도록 합시다. (Transcription, Translation)
Chap11을 통해서는 유전자 발현 조절 과정을 이해할 수 있는데, 기본적으로는 promoter 또는 enhancer 부위의 염기서열에 의한 조절이 가능하고, 동일한 유전적 염기서열이지만 epigenetic modification을 통해 조절이 가능합니다. 조금 생소하고 어려운 개념이긴 하지만 우선 이해해 보도록 노력하고, chap11의 경우는 강의를 통해 다시 한번 설명할 예정입니다.
유전자 발현 과정과 조절 기작
Chap10을 통해 유전자의 기본적인 구조를 이해하고 (Promoter, Enhancer, Intron, Exon etc..) 이들 각 구조의 기능을 정리해 보도록 하고, 유전자의 발현과정을 정리해 보도록 합시다. (Transcription, Translation)
세포는 유전적 지시를 이용하여 단백질을 제조하는 데 두 가지 과정을 거친다. 전사(transcription》는 먼저 특정한 유전자에 해당하는DNA 이중나선의 한 가닥에 대하이 상보적인 RNA 분자를 합성한다. 그 RNA 복사본은 핵으로부터 나와 세포질로 간다. 번역 과정은 RNA에 있는 정보를 이용하이 아미노산서열올 연결 시켜 단백질을 합성한다. 마지막으로 단백질은 기능을 수행하기위한 특정한 3차원 형태로 접혀진다.
RNA는 유전자와 단백질 사이를 연결시키는 역할을 한다. 그림 10.2 에서 묘사한 것처럼 RNA와 DNA는 밀접한 관계를 가지고 있다. RNA 가닥의 염기는 주형 가닥(template strand》이라고 하는 이중 나선의 한 가닥과 상보적이다. RNA 중합효소라는 효소는 RNA 분 자를 만든다. DNA 이중 가닥 중 다른 비주형 가닥은 암호화 가닥 (coding slrand》이라고 불린다.
RNA 의 세 가지 주요 종류는 전령 RNA (messenger RNA),
리보솜 RNA (ribosomal RNA), 운반 RNA(transfcr RNA) 이다(표
W.3》. 다른 종류의 RNA는 특별한 환경에 따라 유전자의 발현(전사
및 번역)을 조절한다.
전령 RNA(mRNA)는 특정 단백질을 구체화하는 정보를 운반한 다. mRNA 일련의 3개의 염기는 코돈(codon)이라고 하는 유전적 암 호를 형성하는데,이 코돈은 특정한 아미노산을 지정한다. 유전자는 길이가 다양하기 때문에 성숙한 mRNA 분자의 길이도 다양하다
mRNA 서열에 들어 있는 정보를 사용하기 위해 세포는 다른 2종 류의 RNA를 필요로 한다. 대부분 리보솜 RNA(rRNA) 분자는 약 100~3,000 뉴클레오티드 길이로 구성된다. 리보솜 RNA들은 단백질 들과 결합하여 리보솜을 형성한다. 리보솜은 서로 다른 여러 가지 단 백질과 RNA 소단위로 이루어진 세포 내 소기관임을 제2장에서 다룬 바 있다. 전반적으로 보았을 때,리보솜은 아미노산을 조립하고 아 미노산을 연결하여 단백질을 형성하는 기능을 수행한다(
rRNA는 아미노산 사이의 펩티드 결합 헝성을 촉매한다. 이러한 효
소의 기능을 가지는 rRNA를 리보자임이라고 한다. 다른 rRNA는 리 보솜과 mRNA가 배열하는 것을 돕는다.
RNA 분자의 세 번째 주요 유형인 운반 RNA(tRNA)는 한쪽 말단
은 mRNA 코돈을 연결시키고 다른 쪽 말단에는 특정한 아미노산을 연결시킨다.
tRNA의 고
리 중 하나는 연속된 3개의 염기를 가지고 안티코돈(anticodon)을 형성하는데 이것은 mRNA 코돈에 상보적이다.
tRNA는 항상 페닐알라닌이라는 아미노산과 결 합한다. 효소는 적절한 안티코돈을 가진 tRNA와 특정 아미노산 사 이에 화학적 결합을 형성하여 부착시킨다
전사인자
세포 종류에 따라 다른 부분의 유전자들이 발현한다. 이것을 수행 하기 위해, 전사인자(transcription factor)라 불리는 단백질들이 모 이고 상호작용하며, 특정한 서열의 DNA에 결합되어 염색체 위의 특 정 부분에 전사를 개시하는 기구를 형성한다. 전사인자는 세포 밖의 호르몬이나 성장인자들과 같은 신호에 반옹하고,RNA 중합효소가 결합하기 위한 주머니를 형성하고 RNA 사슬을 만들기 시작한다. 전 사인자는 그들이 조절해야 할 유전자로 안내하는 결합부위를 가지 고 있다. DNA 결합영역은 나선-회전-나선(helix-tum-helix),’ ‘아연 집게 (zinc fingers); •류신 지퍼 (leucine zipper)’라는 다채로운 이름 을 가지고 있는데 이는 그들의 다양한 형태를 반영한 것이다.
전사의 단계
전사는 3단계로 설명될 수 있다: 개시, 신장 및 종결. 전사 과정은 정 보를 복사하지만 유전 언어인 뉴클레오티드 염기 단계에 머무르게
된다.
전사인자와 RNA 중합효소(RNAP)는 특정한 유전자의 전사를 개 시하기 위해서 DNA의 어느 부위에 결합해야 하는지 '알고' 있다. 전 사 개시에서 전사인자와 RNA 중합효소는 문장의 시작 부분에 있는 대문자와 같이 유전자의 시작을 알리는 특별한 부분인 프로모터
(promoter) 부위에 화학적으로 이끌린다.
그림 10.7은 전사인자의 결합을 간단하게 보여주고 있는데, 이 결 합을 통해 RNA 중합효소를 받아들이게 준비된다.
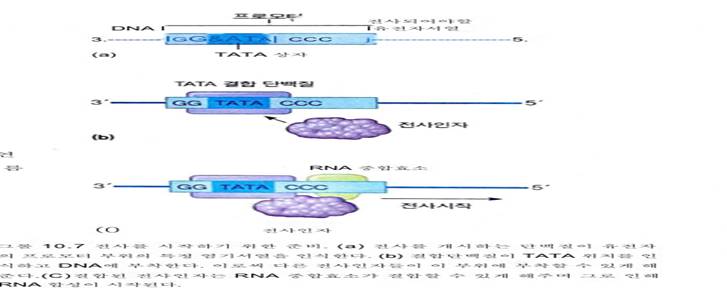

최종적으로 R N A 중합효소가
유전자 시작부위 바로 앞쪽에 결합하면서
복합체를 형성한다. 이러한 요소들의 결합
이 바로 전사의 개시를 알린다.
다음으로 전사의 신장 단계에서는,효소 들이 DNA의 이중나선을 부분적으로 풀 며, 유리된 RNA 뉴클레오티드는 노출된
DNA 주형가닥에 상보적인 염기와 결합한
다(그림 10.2 참조). RNA 중합효소는 주
형가닥의 3'에서 5' 방향으로 DNA 가닥
을 따라 이동하면서, 5'에서 3' 방향으로
주형 DNA가 지정하는 염기서열에 따라
RNA 뉴클레오티드를 붙인다. 주형 DNA
말단 염기서열은 한 문장의 끝부분에 있는
마침혼처럼 유전자의 RNA 암호화 부위가
끝났음을 나타낸다. 이 부위에 이르게 되
면,세 번째 단계인 전사 종결이 일어난다
(그림 10.8). 인간 유전자의 일반적인 전사
속도는 1초당 약 20개 염기이다.
RNA는 일반적으로 유전자의 주형가닥
만을 전사에 이용한다. 하지만 아마도 같
은 염색체에 존재하는 다른 유전자들은 이 중나선의 서로 다른 가닥으로부터 전사될
수 있다. DNA의 암호화 가닥이라 불리는
것은 티민 대신에 우라실로 교체되는 것을 제외하고는 그 서열 자체가 RNA 사슬과
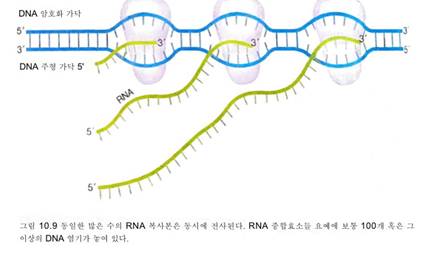
동일하기 때문이다. 몇 개의 …요들은 같
은 DNA 주형 가닥으로부터 동시에 전사
될 수도 있다(그림 10.9). mRNA는 약 10
분에 절반이 분해될 정도로 짧은 수명을 가 지기 때문에. 세포는 필요한 단백질을 유지 하기 위해서 지속적으로 전사해야만 한다.
유전자로부터 전사된 RNA 염기의 서열
을 결정할 때는 DNA 주형 가닥에 대하여
상보적인 RNA 염기를 쓰는데 이때 아데닌 반대편에는 우라실을 쓴다. 에를 들어
D N A 주 형 가 닥 이 3 , - C C T A G C T A
C-5' 서열을 가지면 RNA로 전사된 염기서
열 은 5 ' - G G A U C G A U 를 가 지 며
암호가닥 DNA 서열은 5'-G G ATCG
ATG-3'이다.
세균의 핵은 물리적으로 전사와 번역 두 과정을 분리하지 않기 때문 에,RNA■가 DNA로부터 전사되자마자 단백질로 번역된다 사람과 같은 진핵세포에서는 일단 mRNA가 핵에서 나와 리보솜이 있는 세 포질로 들어온다. mRNA는 복잡한 세포 내에서 번역되기 전에 여러 단개에 걸쳐 변형된다. 인트론이 제거되기 전의 mRNA는 pre-mRNA라고 부른다. 인트 론은 그들 자신의 제거에 관여한다. 그들은 특정 단백질과 결합하여 소형 핵리보뉴클레오단백질(snRNPs) 또는 ‘snurps’를 형성한다.4개의 snurp는 스플라이시오솜(spliceosome)이라고 불리는 구조를 형성하는데,인트론을 잘라내고 엑손을 서로 결합시켜 성숙 mRNA를 형성하여 핵을 빠져나간다.
서로 다른 단백질을 구성하는 기본 단위의 수(20개)는 서로 다른 mRNA를 구성하는 기본 단위의 수(4개)보다 많다. 각 코돈은 1개이상의 mRNA 염기를 가지고 있어야만 한다. 만일 하나의 코돈이 단 하나의 mRNA 염기로 이루어진다면 코돈은 4개의 염기 A, C, G, U에 1개씩의 아미노산을 대응시켜 단지 4개의 아미노산만을 지정할수 있다.
DNA 염기서열이 중복된다면 특정 아미노산은 항상 또 다른 특정 아미노산이 이어져서 단백질 구조를 제한하게 된다. 예를 들어 AUG 는 항상 UG로 시작하는 코돈이 뒤따른다. 이러한 일은 발생하지 않 는다. 그러므로 단백질 암호화하는 DNA 서열은 중복되지 않는다.
비록 유전암호가 중복되지 않을지라도,어떠한 DNA 또는 RNA 염기서열은 '시작’하는 염기에 따라 3개의 다른 번역틀로 읽힐 수 있 다. 그림 10.13에는 방금 논의했던 염기에 대하여 세 가지 유형의 번역틀을 나타냈다. 이것은 3개의 다른 아미노산의 조합을 암호화
한다.
mRNA와 tRNA가 결합하는 리보솜의 부위는 2개의 부위를 가지 는 것으로 묘사된다. 번역이 진행될 때 리보솜에 있는 그 부위의 위치 들은 변하지 않고 남아 있지만,리보솜이 움직임에 따라 mRNA의 다 른 부분을 덮게 된다. PCpcptide’) 부위는 신장하는 아미노산 슬 을 지탱하고,옆에 있는 A ('acceptor) 부위는 사슬에 주가된 다음 아 미노산을 지탱한다. 그림 10.15a에서 단백질이 단지 처음 2개의 아미 노산으로 구성될 때,Met은 P 부위를, Gly는 A 부위를 차지한다.
리보자임 (효소 활성을 가지고 있는 RNA)으로서 기능하는 rRNA 의 도움으로. 아미노산은 펩티드 결합이라고 하는 화학적 결합에 의 해 연결된다. 그러고 나서 첫 번째 tRNA는 같은 종류의 다른 아미노 산과 결합하여 다시 사용되기 위해서 방출된다. 특별한 효소는 | tRNA가 항상 정확한 아미노산과 결합할 수 있도록 한다. 리보솜은 한 코돈씩 mRNA를 따라 내려가게 되어,A 부위에 있던 mRNA는 P 부위에 있게 된다(그림 10.15b). mRNA는 이제 하나의 tRNA가 결합되어 있고,2개의 아미노산은 P 부위에 있는 tRNA로부터 연결되 어 있다. 이것이 폴리펩티드의 시작 부분이다.
그다음에 리보솜은 또 다른 코돈을 향태 mRNA를 따라 내려가고,
세 번째 tRNA가 해당 아미노산(그림 10.15b에서 Cys)을 들여온다. 이 세 번째 아미노산은 다른 2개의 아미노산과 나란히 배열되어,두 번째 아미노산과 펩티드 결합올 형성하여 사슬이 성장하게 된다. 두 번째 아미노산에 붙어 있는 tRNA는 방출되어 재사용된다. 폴리펩티 드는 한 번에 하나의 아미노신을 결합시킨다.
신장 단계는 리보솜의 A 부위가 '정지' 코돈(UGA, UAG,UA시을 가 질 때 멈 추 는 데 . 이 는 정 지 코 돈 에 대 웅 하 는 분 자 가 없 기 때 문시다. 단백질 방출 인자가 폴리펩티드를 방출시키기 시작한다. 마 지막 tRNA가 리보솜올 떠날 때’ 새로운 폴리펩티드가 방출된다(그 림 10.16).
단백질 합성과정은 경제적이다. 하나의 세포는 단지 하나 또는 2개의 유전자 사본으로부터 특정한 단백질 분자를 대량으로 만들 수 있다. 예를 들어 면역체계에시 혈장세포는 초당 2,000개의 동일한 항
체 분자들을 만든다. 이러한 속도로 단백질을 생산하기 위해, RNA,
리보솜, 효소, 그리고 다른 단백질들은 지속적으로 재사용된다. 또
한 전사는 특정한 mRNA 사본들을 많이 만들고, 그림 10.17에서 보 는 바와 같이 각 mRNA는 아마도 수십 개의 리보솜과 결합할 것이
다. 하나의 리보솜이 mRNA를 따라 충분히 멀리 이동하면, 또 다른
리보솜이 결합한다. 이런 방법으로 암호화된 단백질의 많은 사본들
이 같은 _A로부터 만들어진다.
'전공 ~2021 > 생명공학(이론 실험)' 카테고리의 다른 글
| 생물물리화학방법론2-ph (0) | 2022.11.14 |
|---|---|
| 생물물리화학 방법론 -단백질 정제 (0) | 2022.11.14 |
| 암생물학-용어정리 (0) | 2022.11.06 |
| 유전학-세포활동 (0) | 2022.11.06 |
| 유전학 - 개요정리 (0) | 2022.11.06 |
